
2022 год в палеонтологии позвоночных
Палеонтология позвоночных — это раздел палеонтологии, объектом изучения которого являются древние позвоночные (животные, у которых есть позвоночник)[1].
В нижеследующем списке перечислены исследования, связанные с изучением ископаемых остатков позвоночных, опубликованные в рецензируемой литературе в 2022 году.
Исследования общей направленности

- П. А. Безносов и Д. В. Пономарев опубликовали краткий обзор ископаемых позвоночных Республики Коми (Россия)[3].
- Brocklehurst, Ford & Benson применили всеобъемлющий набор филогенетических данных для количественной оценки вариаций темпов и ограничений морфологической эволюции среди амниот карбона и ранней перми[4].
- Werneburg & Abel смоделировали анапсидный череп Captorhinus aguti и, взяв его строение за исходное, предприняли попытку разграничить морфотипы черепов ранних амниот и их ближайших родственников, а также реконструировать челюстную мускулатуру их обладателей[2].
- Marsicano, Gaetano & Irmis опубликовали обзор триасовых сообществ четвероногих, живших на территории нынешней Южной Америки, в котором рассмотрели роль южноамериканских находок для понимания триасовых неморских экосистем Гондваны и Пангеи в целом[5].
- Sues et al. описали скопление остатков четвероногих из среднетриасовых отложений Economy Member формации Вулфвилл (Новая Шотландия, Канада): был обнаружен материал трематозаврида подсемейства Lonchorhynchinae, как минимум двух неописанных таксонов темноспондилов надсемейства Capitosauroidea, дицинодонта клады Kannemeyeriiformes, парарептилий семейства Procolophonidae и нескольких таксонов архозавроморф (в том числе первые остатки Tanystropheus, описанные из восточной части Северной Америки)[6].
- Williams et al. описали остатки микропозвоночных из верхнетриасовых (рэтских) отложений Гарден-Клиф, формация Уэстбери (Англия): хрящевых рыб, костных рыб и рептилий, в том числе кости Pachystropheus, ихтиозавров и фрагмент дуги позвонка, который может принадлежать динозавру; также сообщается об окаменелостях беспозвоночных и копролитах из того же местонахождения[7].
- Maxwell et al. опубликовали исследование остатков крупных позвоночных (рыб и ихтиозавров) из нижнеюрских сланцев Посейдония в Германии, сохранивших свидетельства существования особых биотических сообществ на их трупах[8].
- Moreau et al. описали остатки флоры и фауны из верхнеюрского литографского известняка Кос-Межан (Лозер, Франция), которые были обнаружены в карьерах Дригас (Drigas) и Нивольерс (Nivoliers). Позвоночные известняка представлены несколькими лучепёрыми рыбами, целакантом и клювоголовым Kallimodon cerinensis[9].
- Yu et al. сообщили об обнаружении во Внутренней Монголии нового лагерштетта (Moqi Fossil Lagerstätte), относящегося к нижнему мелу (апту). Предполагается, что фауна Моки (Moqi Fauna) может быть связана с джехольской биотой. Среди фоссилий позвоночных в лагерштетте были найдены остатки бесхвостых земноводных (Genibatrachus baoshanensis), хвостатых земноводных (Nuominerpeton aquilonaris), лучепёрых рыб (Peipiaosteus и, возможно, Jinanichthys / Lycoptera), птиц (Khinganornis hulunbuirensis) и черепах (неописанные вид и род Macrobaenidae)[10]; ранее также кратко сообщалось об обнаружении в тех же отложениях остатков орнитопод и небольших теропод[11][10].
- Dridi описал копролиты плотоядных позвоночных из нижнемеловой формации Айн-эль-Геттар в Тунисе, которые, как предполагается, были произведены костными рыбами, пластиножаберными и крокодиломорфами[12].
- During et al. опубликовали исследование гистологического строения и изотопного состава костей осетровых и веслоносовых рыб из местонахождения Танис (Tanis) в Северной Дакоте (США). Отложения этого местонахождения сформировались в момент импактного события, которое, как предполагается, привело к мел-палеогеновому вымиранию: сейсмические волны вызвали сейши (стоячие волны), в результате чего жившие в этой местности рыбы оказались погребены заживо. По мнению авторов работы, полученные ими данные свидетельствуют о том, что вымирание произошло во время северной весны; это согласуется с выводами аналогичного исследования DePalma et al. (2021[13])[14][15][16].
.png)
- McCurry et al. описали лагерштетт, названный McGraths Flat (Новый Южный Уэльс, Австралия), где были обнаружены хорошо сохранившиеся остатки миоценовой биоты тропических лесов. Помимо микрофоссилий, растений, насекомых и пауков, в отложениях лагерштетта были найдены сравнительно редкие остатки позвоночных: одно контурное перо птицы, вероятно, близкой по размеру к воробью, и более дюжины экземпляров рыб (cf. Retropinnidae и один образец Perciformes indet.). У некоторых особей рыб сохранился желудочно-кишечный тракт вместе с содержимым желудка[17].
- Kjær et al. сообщили об обнаружении фрагментов древнейшей известной экологической ДНК возрастом более двух миллионов лет в формации Кап-Копенгаген (Гренландия). Сравнение секвенированных последовательностей ДНК с уже соотнесёнными с различными таксонами последовательностями позволило воссоздать существовавшую в этой местности экосистему, не имеющую современных аналогов. Согласно результатам исследования, на территории Кап-Копенгаген был бореальный лес с разнообразными флорой и фауной, в том числе позвоночными: мастодонтами, северными оленями, зайцами, грызунами, гусями[18][19][20].
- Murchie et al. реконструировали митохондриальные геномы дикой лошади (Equus caballus), степного зубра (Bison priscus), шерстистого мамонта (Mammuthus primigenius) и белой куропатки (Lagopus lagopus), живших на территории нынешнего центрального Юкона (Канада) около 30 000 лет назад. Образцы экологической ДНК были выделены из ила, обнаруженного в многолетней мерзлоте[21].
- Tilby, Miracle & Barker опубликовали исследование микрофоссилий позвоночных из палеолитической пещеры Шанидар (Иракский Курдистан)[22].
Бесчелюстные и рыбы
Земноводные
Описание опубликовано в 2022 г.
Описание анонсировано в 2022 г.
| Название | Новшество | Авторы | Возраст | Типовая местность | Страна | Примечания | Илл. | ZB |
|---|---|---|---|---|---|---|---|---|
| Albanerpeton ektopistikon[23] | Sp. nov | Carrano et al. | Нижний мел | Формация Кловерли (Cloverly Formation) |
( |
Беспанцирная амфибия. | [1] | |
| Baurubatrachus santosdoroi[24] | Sp. nov | Muzzopappa et al. | Верхний мел | Формация Адамантина (Adamantina Formation) |
( |
Лягушка из подотряда Neobatrachia. | [2] | |
| Buettnererpeton[25] | Gen. et comb. nov | Gee & Kufner | Верхний триас | Группа Докум (Dockum Group) |
( ( |
Темноспондил из семейств метопозаврид; новая комбинация для "Buettneria" bakeri Case, 1931. | 
|
[3] |
| Calyptocephalella sauzalensis[26] | Sp. nov | Nicoli et al. | Миоцен | Формация Эль-Саузаль (El Sauzal Formation) |
( |
Шлемоголовый свистун. | [4] | |
| Chemnitzion[27] | Gen. et sp. nov | Werneburg et al. | Пермь (сакмарско-артинская граница) | Формация Лейкерсдорф (Leukersdorf Formation) |
( |
Темноспондил из семейства Zatracheidae. Включает единственный вид — C. richteri. | 
|
|
| Cretadhefdaa[28] | Gen. et sp. nov | Lemierre & Blackburn | Верхний мел (сеноман) | Формация Дуира (Douira Formation) группы Кем-Кем (Kem Kem Group) | (Драа — Тафилалет) |
Лягушка из подотряда Neobatrachia. Включает единственный вид — C. taouzensis. | ||
| Marmorerpeton wakei[29] | Sp. nov | Jones et al. | Средняя юра | Формация Килмалуаг (Kilmaluag Formation) |
( |
Стволовая саламандра из семейства Karauridae. | ||
| Rhinella xerophylla[30] | Sp. nov | Ponssa et al. | Верхний плиоцен | Формация Укия (Uquía Formation) |
( |
Жаба из ныне живущего рода Rhinella. | ||
| Termonerpeton[31][32] | Gen. et sp. nov | Clack, Smithson & Ruta | Карбон (визей) | Вулканическая формация Батгейт-Хиллс (Bathgate Hills Volcanic Formation) |
( |
Четвероногое неясного систематического положения, возможно, рептилиоморф. Включает единственный вид — T. makrydactylus. | 
|
- Whitney et al. опубликовали исследование гистологии костей Whatcheeria deltae. По мнению авторов работы, полученные ими результаты свидетельствуют о том, что детёныши W. deltae быстро росли и быстро достигали скелетной зрелости[33].
- Calábková, Březina & Madzia опубликовали исследование следовых дорожек сеймуриаморф из пермских (ассельских) отложений Boskovice Basin в Чехии — одних из древнейших следов жизнедеятельности сеймуриаморф в мире. Исследователи интерпретировали описанный ими материал как свидетельствующий о существовании наземных сеймуриаморф, которые были намного крупнее самых больших экземпляров Discosauriscidae, обнаруженных в этой местности, а также о вероятном изменении среды обитания на поздних стадиях онтогенеза сеймуриаморф. Самый большой след обладает высокой сохранностью и включает отпечатки мягких тканей[34].
- Maho & Reisz изучили анатомию зубной системы сеймурий (Seymouria) и схему замены зубов, характерную для представителей этого рода[35].
- Jansen & Marjanović опубликовали исследование микроанатомии костей конечностей и осевого скелета Batropetes palatinus. Палеонтологи пришли к выводу, что животные этого вида вели наземный образ жизни, который включал копание почвы, но не рытьё нор. По их мнению, мощные передние конечности Triadobatrachus massinoti, сочетавшиеся с отсутствием способности прыгать, могли быть прежней адаптацией к копанию, которая в дальнейшем сделала возможными прыжки бесхвостых земноводных в результате экзаптации[36].
- Arbez, Atkins & Maddin изучили образец темноспондила из каменноугольной формации Джоггинс (Новая Шотландия, Канада), который первоначально был отнесён к виду Dendrysekos helogenes. Исследователи пришли к выводу, что Dendrysekos, скорее всего, является младшим синонимом Dendrerpeton[37].
- Herbst, Manafzadeh & Hutchinson опубликовали исследование, обосновавшее гипотезу о том, что конфигурация задних конечностей Eryops megacephalus была подобна таковой у саламандр[38].
- Hart, Campione & McCurry опубликовали исследование, направленное на оценку массы тела Eryops megacephalus и Paracyclotosaurus davidi[39].
- А. В. Ульяхин опубликовал исследование, направленное на выявление и уточнение палеоэкологических особенностей темноспондила платиопозавра (Platyoposaurus) из средней перми Башкортостана (Россия)[40].
- Gee & Sidor описали остатки крупных темноспондилов клад Capitosauria и Plagiosauridae из среднетриасовой формации Фремоу (Антарктида). Новые и обнаруженные прежде в формации образцы были интерпретированы как свидетельствующие о родстве Capitosauria и Trematosauria[41].
- Witzmann & Schoch опубликовали переописание Platycepsion wilkinsoni. Палеонтологи интерпретировали остатки этого брахиопида как принадлежавшие настоящей личинке, что подтверждает присутствие личиночной стадии развития в онтогенезе стереоспондильных[42].
- Schoch & Mujal описали новые ископаемые остатки Trematolestes hagdorni, благодаря изучению которых были получены дополнительные данные об онтогенезе и анатомии взрослых особей этого темноспондила[43].
- Kalita et al. опубликовали исследование компактности костей в плечевом поясе Metoposaurus krasiejowensis. По мнению палеонтологов, полученные ими результаты указывают на то, что компактный передний конец межключичных костей вместе с тяжёлым черепом этого метопозаврида мог действовать как балласт для контроля гидростатической плавучести путём смещения центра масс вперёд[44].
- Weryński & Kędzierski опубликовали исследование микроструктуры зубов Metoposaurus krasiejowensis. Авторы работы сообщили об обнаружении возможных приспособлений к хищничеству, а также о свидетельствах сезонного влияния на гистологию зубов[45].
- Surmik et al. (2022) описали патологический интерцентр позвонка Metoposaurus krasiejowensis из местонахождения Красеюв в Польше. Палеонтологи диагностировали экземпляр как поражённый остеосаркомой. Это первый образец мезозойского позвоночного, не принадлежащего к амниотам, который содержит недвусмысленные свидетельства первичной злокачественной опухоли кости[46].
- Schoch & Sues переописали темноспондила Parioxys ferricolus из нижней перми Техаса (США)[47].
- Schoch опубликовал исследование филогенетических взаимоотношений представителей клады Amphibamiformes[48].
- Macaluso et al. опубликовали исследование биогеографической истории палеарктических хвостатых земноводных, в котором провели ревизию ископаемых остатков представителей этой группы, обнаруженных в Палеарктике[49].
- Jia, Li & Gao опубликовали исследование нёбных костей современных и вымерших саламандр (тотальная группа Caudata). Авторы работы пришли к выводу, что морфология нёба является надежным показателем для реконструкции палеобиологии ранних саламандр[50].
- Gardner описал неполную зубную кость саламандры из верхнемеловой (маастрихтской) формации Ланс (Вайоминг, США), предположительно принадлежащую новому виду и роду семейства Batrachosauroididae[51].
- Ledesma, Ayala & Kemp провели морфометрический анализ позвонков амбистом (Ambystoma), чтобы определить, насколько надёжно могут быть идентифицированы ископаемые остатки хвостатых земноводных[52].
- Skutschas et al. (2022) описали фрагментарный туловищный позвонок кроновой саламандры из среднеюрской (батской) москворецкой свиты в Московской области России. Это первый материал кроновой саламандры, обнаруженный в средней юре Восточной Европы. Если отнесение позвонка к Salamandroidea верно, то это древнейшие известные остатки представителя данного надсемейства[53].
- Barcelos et al. опубликовали исследование филогении современных и вымерших Ceratophryidae[54].
- Guevara et al. описали остатки жабы (Bufonidae) cf. Rhinella из миоценовых (серравальских) отложений формации Кура-Маллин в Чили; это самая южная окаменелость жабы, обнаруженная в миоцене Южной Америки[55].
Завропсиды
| Название | Новшество | Авторы | Возраст | Типовая местность | Страна | Примечания | Илл. | ZB |
|---|---|---|---|---|---|---|---|---|
| Baisesaurus[56] | Gen. et sp. nov | Ren et al. | Нижний триас (оленёк) | Формация Луолоу (Luolou Formation) |
Базальный представитель Ichthyosauromorpha. Включает единственный вид — B. robustus. | |||
| Magnipterygius[57] | Gen. et sp. nov | Maisch & Matzke | Нижняя юра | Формация Посидониеншифер (Posidonienschiefer Formation) | Предварительно отнесён к семейству Stenopterygiidae. Включает единственный вид — M. huenei. | |||
| Nannopterygius mikhailovi[58] | Sp. nov | Yakupova & Akhmedenov | Верхняя юра | (Западно-Казахстанская область) |
Ихтиозавр из семейства Ophthalmosauridae. | |||
| Nannopterygius yakimenkae[58] | Sp. nov | Yakupova & Akhmedenov | Верхняя юра | (Западно-Казахстанская область) |
Ихтиозавр из семейства Ophthalmosauridae. |
- Nakajima et al. описали позвонки, ребро и плечевую кость ихтиоптеригий, обнаруженные в отложениях нижнетриасовой житковской свиты (остров Русский, Приморский край, Россия). Длина ящера, которому принадлежала плечевая кость, была оценена в 5 метров, что делает его одной из самых крупных известных на сегодняшний день морских рептилий, живших в начале спатского подвека оленёкского века[59].
- Holm, Delsett & Alsen предприняли попытку использовать пропорции тел позвонков ихтиозавров из верхнеюрской формации Бернбьерг (Гренландия), чтобы определить, какое место в позвоночнике занимали повреждённые позвонки[60].
- Roberts, Engelschiøn & Hurum описали новый экземпляр Phalarodon из среднетриасовых (ладинских) отложений Blanknuten Member формации Ботнехейя (Botneheia Formation) на архипелаге Шпицберген в Норвегии. Это первый трёхмерный череп Mixosauridae, обнаруженный в этой формации[61].
- Kelley et al. (2022) изучили скопление скелетов Shonisaurus popularis из верхнетриасовой формации Лунинг (Невада, США), в котором были обнаружены остатки взрослых шонизавров, а также нескольких эмбрионов или новорожденных детенышей. Палеонтологи интерпретировали скопление скелетов как «ясли» ихтиозавров и предположили, что ради размножения шонизавры мигрировали в низкопродуктивные умеренно глубокие воды[62][63].
- Lukeneder et al. описали первые два экземпляра ихтиозавров (клада Ophthalmosauria), обнаруженные в нижнемеловых отложениях Австрии (Северные Известняковые Альпы; валанжин — готерив). Исследователи пришли к выводу, что готеривские ихтиозавры существенно отличались от тех, что жили в титонском и берриасском веках, и предположили существование некоего значимого события в эволюционной истории ихтиозавров, которое должно было произойти во времена валанжинского века[64].
- Eriksson et al. провели обзор исследований ихтиозавров, опубликованных за последние 300 лет, фокусируясь на тех работах, что касаются мягких тканей. В частности, были проанализированы выдвигавшиеся в разное время предположения о различных аспектах биологии ихтиозавров, таких как окраска и терморегуляция. При этом палеонтологи изучили исключительно хорошо сохранившийся образец Stenopterygius из нижнеюрских сланцев Посейдония (Германия), уточнив детали анатомии и физиологии ихтиозавров. В рамках исследования была создана детальная трёхмерная реконструкция стеноптеригия в натуральную величину, которую авторы работы называют наиболее современной с научной точки зрения реконструкцией ихтиозавра, доступной в настоящее время[65][66].
- Miedema & Maxwell опубликовали исследование пренатальных и постнатальных онтогенетических изменений в черепе Stenopterygius quadriscissus[67].
- Delsett et al. описали практически полный скелет Aegirosaurus sp. и изолированный хвост неопределимого ихтиозавра, обнаруженные в зольнхофенскм известняке на юге Германии (формация Айхштетт; титон, верхняя юра). Наблюдения в ультрафиолетовом свете подтвердили наличие ископаемых остатков кожи у обоих экземпляров. Кроме того, у первого образца были обнаружены мягкие ткани, интерпретированные как окаменевший жировоск, и неопределённые ткани в области живота[68][69].
- Serafini et al. провели ревизию материала кончиков челюстей Platypterygiinae из мела Северных Апеннин (Италия) и обнаружили каналы внутри костей, которые могли предназначаться для тройничного нерва[70].
- Laboury et al. опубликовали исследование анатомии и филогенетического положения «Ichthyosaurus» zetlandicus, в котором обосновали принадлежность этого вида к роду Temnodontosaurus[71].
| Название | Новшество | Авторы | Возраст | Типовая местность | Страна | Примечания | Илл. | ZB |
|---|---|---|---|---|---|---|---|---|
| Eardasaurus[72] | Gen. et sp. nov | Ketchum & Benson | Средняя юра (келловей) | Оксфорд-Клей (Oxford Clay) | ( |
Плезиозавр из семейства плиозаврид. Включает единственный вид — E. powelli. | 
|
|
| Honghesaurus[73] | Gen. et sp. nov | Xu et al. | Средний триас (анизий) | Формация Гуаньлин (Guanling Formation) |
(Юньнань) |
Представитель семейства Pachypleurosauridae. Включает единственный вид — H. longicaudalis. | 
|
|
| Nothosaurus luopingensis[74] | Sp. nov | Shang, Li & Wang | Средний триас (анизий) | Формация Гуаньлин | (Юньнань) |
Представитель семейства Nothosauridae отряда Nothosauroidea. | ||
| Plesioelasmosaurus[75] | Gen. et sp. nov | Schumacher & Everhart | Верхний мел (сеноман) | Известняк Гринхорн (Greenhorn Limestone) |
( |
Плезиозавр из семейства эласмозаврид. Включает единственный вид — P. walkeri. | 
|
|
| Prosantosaurus[76] | Gen. et sp. nov | Klein et al. | Средний триас (ладин) | Формация Просанто (Prosanto Formation) | ( |
Представитель подотряда Pachypleurosauria. Включает единственный вид — P. scheffoldi. | 
|
|
| Serpentisuchops[77][78] | Gen. et sp. nov | Persons, Street & Kelley | Верхний мел (маастрихт) | Пьер-Шейл (Pierre Shale) |
( |
Плезиозавр из семейства поликотилид. Включает единственный вид — S. pfisterae. | [5] |
- Klein et al. описали две зубные кости плакодонтов из нижнего триаса (мушелькалка) Нидерландов и Германии, предварительно отнесённые к Placodus cf. gigas. Исследователи предприняли попытку выявить влияние онтогенетических изменений на морфологию зубной кости плакодонтов[79].
- Madzia, Sachs & Klug переописали типовой материал плиозаврида Ischyrodon meriani; авторы работы пришли к выводу, что данный вид лучше рассматривать как nomen dubium[80].
- Krahl et al. опубликовали исследование силы и функции мышц конечностей плезиозавра Cryptoclidus eurymerus[81].
- Brum et al. опубликовали исследование морфометрии шейных позвонков антарктических эласмозаврид в контексте их филогенеза и онтогенеза[82].
- Wiemann et al. применили рамановскую и инфракрасную спектроскопию с преобразованием Фурье для количественной оценки прижизненного накопления конечных продуктов перекисного окисления липидов (advanced lipoxidation end-products, ALE) в костях современных и вымерших рептилий. Одним из выводов исследования является независимое приобретение плезиозаврами скорости метаболизма, соответствующей эндотермии[83].
| Название | Новшество | Авторы | Возраст | Типовая местность | Страна | Примечания | Илл. | ZB |
|---|---|---|---|---|---|---|---|---|
| Navajosphenodon[84] | Gen. et sp. nov | Simões, Kinney-Broderick & Pierce | Средняя юра (синемюр — плинсбах) | Формация Кайента (Kayenta Formation) |
( |
Клювоголовое клады Sphenodontinae. Включает единственный вид — N. sani. | ||
| Opisthiamimus[85] | Gen. et sp. nov | DeMar, Jones & Carrano | Верхняя юра (киммеридж — титон) | Формация Моррисон (Morrison Formation) |
Базальное клювоголовое. Включает единственный вид — O. gregori. | [6] |
Описание опубликовано в 2022 г.
Описание было анонсировано в 2021 г., но публикация его финальной версии состоялась в 2022 г.
Описание анонсировано в 2022 г.
| Название | Новшество | Авторы | Возраст | Типовая местность | Страна | Примечания | Илл. | ZB |
|---|---|---|---|---|---|---|---|---|
| Abronia cuyama[86] | Sp. nov | Scarpetta & Ledesma | Миоцен | Формация Калиенте (Caliente Formation) |
( |
Представитель рецентного рода аброний из семейства веретеницевых. | ||
| Archaeovaranus[87] | Gen. et sp. nov | Dong et al. | Нижний эоцен | Формация Юйхуандин (Yuhuangding Formation) |
(Хубэй) |
Представитель варановых. Включает единственный вид — A. lii. | ||
| Blutwurstia[88] | Gen. et sp. nov | Smith, Bhullar & Bloch | Нижний эоцен | Формация Уиллвуд (Willwood Formation) |
( |
Представитель ксенозавров. Включает единственный вид — B. oliviae. | ||
| Cryptovaranoides[89] | Gen. et sp. nov | Whiteside, Chambi-Trowell & Benton | Верхний триас (рэт) | Cromhall Quarry, Тортворт | ( |
Ранний представитель кроновой группы чешуйчатых, предположительно, член клады Neoanguimorpha. Включает единственный вид — C. microlanius. | ||
| Dollogekko[90] | Gen. et sp. nov | Čerňanský et al. | Нижний эоцен | Формация Тинен (Tienen Formation) |
Представитель гекконообразных. Включает единственный вид — D. dormaalensis. | |||
| Eoscincus[91] | Gen. et sp. nov | Brownstein et al. | Верхняя юра (титон) | Формация Моррисон (Morrison Formation) |
( |
Представитель Pan-Scincoidea. Включает единственный вид — E. ornatus. | ||
| Geiseleptes[92] | Gen. et sp. nov | Villa, Wings & Rabi | Эоцен | Гейзельтальский буроугольный бассейн | ( |
Гекконообразная ящерица из семейства Sphaerodactylidae. Включает единственный вид — G. delfinoi. | [7] | |
| Katariana[93] | Nom. nov | Deshmukh et al. | Нижний палеоцен (дат) | Формация Санта-Люсия (Santa Lucía Formation) |
Змея из клады Macrostomata, близкая к Caenophidia. Замена названия Kataria Scanferla et al., 2013. | |||
| Microteras[91] | Gen. et sp. nov | Brownstein et al. | Верхняя юра (титон) | Формация Моррисон | ( |
Представитель Pan-Scincoidea. Включает единственный вид — M. borealis. | ||
| Moqisaurus[94] | Gen. et sp. nov | Dong, Wang, & Evans | Нижний мел (апт) | (Внутренняя Монголия) |
Базальное чешуйчатое, предположительно близкое к Liushusaurus. Включает единственный вид — M. pulchrum. | |||
| Morohasaurus[95] | Gen. et sp. nov | Ikeda et al. | Нижний мел | Формация Охьямашимо (Ohyamashimo Formation) |
Веретеницеобразная ящерица из клады Monstersauria. Включает единственный вид — M. kamitakiensis. | |||
| Powellophis[96] | Gen. et sp. nov | Garberoglio, Triviño & Albino | Средний палеоцен — верхний палеоцен | Формация Меалла (Mealla Formation) |
( |
Змея из семейства Madtsoiidae. Включает единственный вид — P. andina. | [8] | |
| Psammophis odysseus[97] | Sp. nov | Szyndlar & Georgalis | Верхний миоцен | ( |
Представитель рецентного рода песчаных змей из семейства Lamprophiidae. | |||
| Retinosaurus[98] | Gen. et sp. nov | Čerňanský et al. | Нижний мел (апт) | Бирманский янтарь (Hkamti amber) |
Сцинкообразная ящерица, возможно, представитель Pan-Xantusiidae. Включает единственный вид — R. hkamtiensis. | 
|
||
| Smithosaurus[99] | Gen. et sp. nov | Vasilyan et al. | Миоцен | Представитель подсемейства Anguinae семейства веретеницевых. Включает единственный вид — S. echzellensis. | [9] | |||
| Thalassotitan[100] | Gen. et sp. nov | Longrich et al. | Верхний мел (маастрихт) | Бассейн Улед-Абдун (Ouled Abdoun Basin) |
(Бени-Меллаль — Хенифра) |
Мозазавр из трибы Prognathodontini. Включает единственный вид — T. atrox. | 
|
- Woolley et al. пришли к выводу, что, несмотря на неполноту палеонтологической летописи чешуйчатых, она содержит надёжную филогенетическую информацию[101].
- Yang et al. описали первые известные ископаемые остатки сцинкообразных ящериц из верхнемеловой (кампанской) формации Ненцзян (Nenjiang Formation) в китайской провинции Гирин[102].
- Vullo et al. переописали фрагмент нижней челюсти, который, как считалось ранее, происходит из нижнемеловых отложений Кем-Кем-Бедс в Марокко и принадлежит единственному известному виду игуанообразных из мезозоя Африки — Jeddaherdan aleadonta. Палеонтологи пришли к выводу, что на самом деле материал имеет четвертичный возраст и принадлежит шипохвосту неопределённого вида (Uromastyx sp.)[103].
- Syromyatnikova & Aranda описали первые известные ископаемые остатки веретеницеобразной ящерицы Diploglossus с Кубы, обнаруженные в верхнем плейстоцене пещеры Эль-Аброн (El Abrón Cave). Получены первые данные о морфологии зубов и челюстей плейстоценовых Diploglossus с Кубы[104].
- Syromyatnikova, Klembara & Redkozubov описали ископаемые остатки веретеницевых Ophisaurus fejfari и Ophisaurus spinari из нижнего плиоцена Молдовы. Это первый материал рода Ophisaurus из плиоцена Восточной Европы, а также первый описанный материал O. fejfari из плиоценовых отложений вообще[105].
- Augé et al. переописали известный материал варана Saniwa, а также описали новый материал этого вида из эоцена Бельгии и Франции[106].
- Villa & Delfino описали позвонок варана (Varanus), предположительно, обнаруженный в формации Чинджи (Chinji Formation). Это первые остатки ящерицы из неогеновых отложений Пакистана. Открытие согласуется с более ранними предположениями о тёплом климате на территории Сивалика в среднем миоцене — начале позднего миоцена[107].
- Cross et al. опубликовали исследование диверсификации стратегий питания и передвижения мозазавроидей[108].
- MacLaren et al. опубликовали исследование морфофункционального разнообразия мозазаврид перед мел-палеогеновым вымиранием. Палеонтологи пришли к выводу, что смена таксономического состава в сообществах мозазаврид при переходе от кампанских сообществ к маастрихтским сопровождалась глобальным увеличением морфофункционального разнообразия. При этом, согласно исследованию, морфофункциональное разнообразие мозазаврид снижалось во многих позднемаастрихтских провинциальных сообществах вплоть до мел-палеогенового вымирания[109].
- Bennion et al. опубликовали исследование эволюции черепа мозазаврид и ранних китообразных в течение первых 20 миллионов лет их эволюционной истории, в котором проверили возможные случаи экоморфологической конвергенции черепа и зубов у этих групп[110].
- Rempert et al. опубликовали первое сообщение об остатках Mosasaurus hoffmannii, представленных коронками зубов, из верхнемеловых (маастрихтских) фосфатных отложений Ouled Abdoun Basin в Марокко[111].
- Zaher et al. изучили череп меловой змеи Sanajeh indicus и рассмотрели его в контексте происхождения характерного для змей способа кормления с широко раскрываемыми челюстями[112].
- Chuliver, Scanferla & Smith описали окаменелость беременной самки земляного удава Messelophis variatus из эоцена карьера Мессель (Германия), в утробе которой были обнаружены остатки как минимум двух детёнышей. Это первый скелет беременной змеи, обнаруженный палеонтологами. Тот факт, что Messelophis жил в «парниковом» климате, поставил под вопрос общепризнанный сценарий возникновения живорождения у чешуйчатых, согласно которому возможность откладывать яйца ограничивал холодный климат. Исследователи выдвинули несколько гипотез о том, какие преимущества могло обеспечить живорождение тропической змее: так, живорождение могло уберегать яйца от затопления или обеспечивать появление на свет немногочисленного, но относительно крупного потомства, способного выдержать конкуренцию в условиях большого количества змей других видов[113][114].
- El-Hares et al. описали позвонки безногих чешуйчатых из верхнеэоценовых отложений Египта: первый материал двуходок, описанный из палеогена Египта, а также остатки змей Renenutet enmerwer, включая первые известные хвостовые позвонки вида, и cf. Procerophis. Присутствие cf. Procerophis и Renenutet в Египте дополняет свидетельства обмена наземной фауной между Азией и Северной Африкой в течение раннего/среднего эоцена вдоль южной окраины моря Тетис[115].
Описание опубликовано в 2022 г.
Описание анонсировано в 2022 г.
| Название | Новшество | Авторы | Возраст | Типовая местность | Страна | Примечания | Илл. | ZB |
|---|---|---|---|---|---|---|---|---|
| Annemys variabilis[116] | Sp. nov | Obraztsova et al. | Средняя юра (бат) | Итатская свита | ( |
Представитель семейства Xinjiangchelyidae. | [10] | |
| Appalachemys[117] | Gen. et sp. nov | Gentry, Kiernan & Parham | Верхний мел (сантон — кампан) | ( |
Представитель грады Macrobaenidae. Включает единственный вид — A. ebersolei. | |||
| Calissounemys[118] | Gen. et sp. nov | Tong et al. | Верхний мел (кампан) | ( |
Представитель семейства Compsemydidae. Включает единственный вид — C. matheroni. | |||
| Chelonoidis gersoni[119] | Sp. nov | Viñola-López & Almonte | Четвертичный (верхний плейстоцен — голоцен) | Американская сухопутная черепаха. | ||||
| Chelonoidis meridiana[120] | Sp. nov | Vlachos, de la Fuente & Sterli | Миоцен | ( |
Американская сухопутная черепаха. | [11] | ||
| Chelydropsis aubasi[121] | Sp. nov | Joyce, Landréat & Rollot | Эоцен (бартон) | ( |
Представитель стволовой группы каймановых черепах (Pan-Chelydridae). | |||
| Dortoka vremiri[122] | Sp. nov | Augustin et al. | Верхний мел (маастрихт) | Формация Санпетру (Sânpetru Formation) |
( |
Представитель стволовой группы бокошейных черепах из вымершего семейства Dortokidae. | ||
| Edowa[123] | Gen. et sp. nov | Adrian et al. | Верхний мел (турон) | Формация Морено-Хилл (Moreno Hill Formation) |
( |
Представитель семейства Baenidae клады Paracryptodira. Включает единственный вид — E. zuniensis. | [12] | |
| Gestemys[124] | Gen. et sp. nov | De la Fuente et al. | Эоцен | Формация Геште (Geste Formation) |
( |
Бокошейная черепаха семейства Podocnemididae. Включает единственный вид — G. powelli. | ||
| Hesperotestudo (Caudochelys) harrisi[125] | Sp. nov | Lichtig & Lucas | Миоцен (Late Barstovian) | Формация Тесук (Tesuque Formation) |
( |
Сухопутная черепаха. | ||
| Hutchemys walkerorum[126] | Sp. nov | Jasinski et al. | Верхний мел (маастрихт) | Формация Хелл-Крик (Hell Creek Formation) |
( |
Трёхкоготная черепаха (под)семейства Plastomeninae/Plastomenidae. | ||
| Jimemys[127] | Gen. et sp. nov | Edgar et al. | Верхний мел (сантон — кампан) | Формация Милк-Ривер (Milk River Formation) |
( |
Трёхкоготная черепаха (под)семейства Plastomeninae/Plastomenidae. Включает единственный вид — J. glaebosus. | ||
| Leiochelys[128] | Gen. et sp. nov | Brinkman et al. | Верхний мел (маастрихт) | Формация Френчмен (Frenchman Formation) |
( |
Представитель стволовой группы иловых черепах (Pan-Kinosternidae). Включает единственный вид — L. tokaryki. | ||
| Leviathanochelys[129][130] | Gen. et sp. nov | Castillo-Visa et al. | Верхний мел (кампан) | Формация Перлес (Perles Formation) |
( |
Крупная морская черепаха. Включает единственный вид — L. aenigmatica. | 
|
|
| Mokelemys[131] | Gen. et sp. nov | Pérez-García | Плиоцен | Представитель трибы Erymnochelyini семейства Podocnemididae. Включает единственный вид — M. mbembe. | ||||
| Prochelidella palomoi[132] | Sp. nov | Maniel, de la Fuente & Filippi | Верхний мел (кампан) | Формация Анаклето (Anacleto Formation) |
( |
Змеиношеяя черепаха. | ||
| Solitudo[133] | Gen. et comb. et sp. nov | Valenti et al. | Плиоцен — плейстоцен | Сухопутная черепаха. Включает три вида:
|
[13] | |||
| aff. "Stylemys" gisellae[134] | Sp. nov | Carbot-Chanona et al. | Олигоцен (Arikareean) | Формация Чилапа (Chilapa Formation) |
Представитель стволовой группы сухопутных черепах. | |||
| Testudo lohanica[135] | Sp. nov | Pérez-García et al. | Верхний миоцен | Crețești 1 | ( |
Вид европейских сухопутных черепах. | ||
| Testudoolithus tuberi (оовид)[136] | Oosp. nov | Ferguson & Tapanila | Верхний миоцен | Формация Кайпаровиц (Kaiparowits Formation) |
( |
Кладка черепашьих яиц с уникальными признаками скорлупы. | ||
| Tongemys[137] | Gen. et sp. nov | Joyce et al. | Нижний мел (берриас) | Группа Пурбек (Purbeck Group) |
( |
Представитель семейства Compsemydidae. Включает единственный вид — T. enigmatica. | [14] |
- Ascarrunz & Sánchez-Villagra опубликовали обзор эволюционной истории и онтогенетического развития щитков карапакса современных и ископаемых черепах[138].
- Rollot et al. опубликовали исследование анатомии черепа Lakotemys australodakotensis из нижнего мела Южной Дакоты (США)[139].
- Xu et al. описали яйца черепах из верхнемеловой формации Гаогоу (Хэнань, Китай). Путём кристаллографического сравнения обнаруженных яиц с яйцами черепах из провинции Чжэцзян и Монтаны были выявлены детали превращения арагонита в кальцит в яичной скорлупе черепах[140].
- Danilov et al. описали новый материал стволовых Chelonioidea из верхнемеловых (кампанских) отложений местонахождения Белое озеро (рыбушкинская свита; Саратовская область, Россия). Среди прочих остатков были описаны образцы Protostega gigas, предполагаемый размер которых соответствует крупнейшим экземплярам из Северной Америки; это первые фоссилии данного вида, обнаруженные за пределами Северной Америки[141].
| Название | Новшество | Авторы | Возраст | Типовая местность | Страна | Примечания | Илл. | ZB |
|---|---|---|---|---|---|---|---|---|
| Puercosuchus[142] | Gen. et sp. nov | Marsh et al. | Верхний триас (норий) | Формация Чинле (Chinle Formation) |
( |
Представитель семейства Azendohsauridae клады Allokotosauria. Включает единственный вид — P. traverorum. | [15] | |
| Stenoscelida[143] | Gen. et sp. nov | Müller, Garcia & Fonseca | Верхний триас (верхний карний / нижний норий) | Формация Санта-Мария (Santa Maria Formation) |
( |
Представитель семейства Proterochampsidae клады Proterochampsia. Включает единственный вид — S. aurantiacus. | [16] | |
| Vigilosaurus[144] | Gen. et sp. nov | Chen & Liu | Верхняя пермь (чансин) | Формация Годикенг (Guodikeng Formation) |
(Синьцзян) |
Архозавриформ, предположительно, из клады Proterochampsia. Включает единственный вид — V. gaochangensis. |
- Pradelli, Leardi & Ezcurra опубликовали исследование размеров тела архозавроморф в течение первых 90 млн лет их эволюции[145].
- Bestwick et al. опубликовали исследование эволюции относительных размеров черепа у палеозойских и мезозойских архозавроморф. Исследователи заключили, что относительные размеры черепов эритрозухид и тероподовых динозавров отличаются друг от друга и что непропорционально большие черепа эритрозухид уникальны среди всех наземных архозавроморф[146].
- Scartezini & Soares описали новые экземпляры гиперодапедонов (Hyperodapedon) из Бразилии с верхушечно зубчатыми коронками зубов, представляющими новый морфотип зубов верхней челюсти и первый случай обнаружения зубчатых зубов у ринхозавров. Было проведено исследование, направленное на то, чтобы определить, является ли морфология верхнечелюстных коронок зубов ринхозавров таксономически информативной или она отражает онтогенез[147].
- Chambi-Trowell et al. опубликовали исследование анатомии и филогенетического положения архозавроморф Tricuspisaurus thomasi и Variodens inopinatus[148].
- Sengupta & Bandyopadhyay опубликовали исследование анатомии и филогенетического положения архозавроморфа Shringasaurus indicus[149].
- Wilson et al. изучили черепную коробку (нейрокраниум) Trilophosaurus buettneri, после чего рассмотрели полученные данные в контексте эволюции нейрокраниума ранних архозавроморф[150].
- Fabbri & Bhullar описали эндокран эупаркерии (Euparkeria capensis) и интерпретировали морфологию её мозга как в целом сходную с таковой у фитозавров, крокодилов и ранних динозавров[151].
- Ezcurra & Sues опубликовали исследование остеологии и филогенетического положения Sphodrosaurus pennsylvanicus, в котором обосновали принадлежность этого таксона к архозавриформам семейства Doswelliidae[152].
Описание анонсировано в 2022 г.
| Название | Новшество | Авторы | Возраст | Типовая местность | Страна | Примечания | Илл. | ZB |
|---|---|---|---|---|---|---|---|---|
| Belebey shumovi[153] | Sp. nov | Bulanov et al. | Средняя пермь (казанский ярус) | ( |
Парарептилия семейства болозаврид. | |||
| Champsosaurus norelli[154] | Sp. nov | Brownstein | Верхний палеоцен (тиффаний) | Формация Полкэт-Бенч (Polecat Bench Formation) |
( |
Вид хампсозавров. | 
|
|
| Kosmodraco[154] | Gen. et comb. et sp. nov | Brownstein | Палеоцен | ( |
Представитель хористодер из семейства Simoedosauridae. Включает типовой вид "Simoedosaurus" dakotensis Erickson, 1987 и новый вид K. magnicornis. | 
|
[17] | |
| Maiothisavros[155] | Gen. et sp. nov | Mooney et al. | Нижняя пермь (артин) | Ричард-Спёр (Richards Spur) |
( |
Ранний представитель клады Neodiapsida. Включает единственный вид — M. dianeae. | ||
| Nagini[156] | Gen. et sp. nov | Mann, Pardo & Maddin | Верхний карбон | Мазон-Крик (Mazon Creek) |
( |
Представитель семейства Molgophidae клады Recumbirostra, рассматриваемой авторами описания в составе завропсид. Включает единственный вид — N. mazonense. | ||
| Pomolispondylus[157] | Gen. et sp. nov | Cheng et al. | Нижний триас (оленёк) | Формация Цзялинцзян (Jialingjiang Formation) |
(Хубэй) |
Представитель новой группы Saurosphargiformes, близкий к семейству Saurosphargidae. Включает единственный вид — P. biani. | ||
| Quasicolognathus[158] | Gen. et sp. nov | Sues, Kligman & Schoch | Средний триас (ладин) | Формация Эрфурт (Erfurt Formation) |
Представитель нового семейства Colognathidae. Включает единственный вид — Q. eothen. |
- Callender-Crowe & Sansom сравнили признаки скелета и мягких тканей у современных завропсид и пришли к выводу, что остеология в большей степени, чем мягкие ткани, соответствует данным молекулярной генетики. Таким образом, работа косвенно поддерживает теоретическую возможность воссоздания сравнительно точной филогении вымерших завропсид, основанной на остеологических данных[159].
- Núñez Demarco, Ferigolo & Piñeiro провели исследование пропорций черепа и костей конечностей более чем сотни образцов мезозавров из Африки, Бразилии и Уругвая с целью определить, существовали ли в их онтогенезе статистически значимые морфологические различия. Исследователи рассмотрели выявленный изометрический рост с точки зрения филогении и паттернов роста ранних амниот[160].
- Verrière & Fröbisch обнаружили свидетельства отногенетического сдвига ниши у мезозавров. Исследователи поддержали интерпретацию Mesosaurus tenuidens как единственного валидного вида в семействе Mesosauridae; по их мнению, экземпляры, отнесённые к Stereosternum tumidum и Brazilosaurus sanpauloensis, представляют собой остатки неполовозрелых M. tenuidens или неполные образцы этого вида[161].
- Jenkins & Bhullar опубликовали исследование по имплантации, замене и прикреплению зубов у проколофономорфа Scoloparia glyphanodon[162].
- Van den Brandt et al. опубликовали исследование таксономии, филогении и стратиграфического распространения парейазавров из пермских (гваделупских) отложений бассейна Кару в ЮАР[163].
- Sidor et al. описали новый материал капторинида Moradisaurus grandis (два неполных черепа молодых особей) из пермской формации Моради в Нигере[164].
- Scheyer et al. описали крупное ребро, несущее остеодерму, которое было обнаружено в верхнетриасовой формации Кессен (Швейцария). Исследователи предварительно определили экземпляр как принадлежащий представителю cf. Saurosphargidae, отметив сходство с родом Largocephalosaurus; такая атрибуция продлевает время существования Saurosphargidae примерно на 35 миллионов лет в поздний триас[165].
- Qin, Yi & Gao описали неоморфное окостенение между теменной, квадратной и чешуйчатой костями хористодер Coeruleodraco jurassicus и Philydrosaurus proseilus[166].
- Wang et al. описали новый экземпляр хористодеры Hyphalosaurus с хорошо сохранившимися покровными остатками, который был обнаружен в нижнемеловых отложениях формации Исянь (Ляонин, Китай)[167].
Синапсиды
- Marugán-Lobón, Gómez-Recio & Nebreda опубликовали исследование морфологического разнообразия черепов синапсид[168].
- Brent, Buchholtz & Mansfield опубликовали исследование происхождения и эволюционной истории грудины млекопитающих[169].
Описание анонсировано в 2022 г.
Выпуск отнесён к 2022 г., но электронная версия опубликована в 2023 г.
| Название | Новшество | Авторы | Возраст | Типовая местность | Страна | Примечания | Илл. | ZB |
|---|---|---|---|---|---|---|---|---|
| Bienotheroides xingshanensis[170] | Sp. nov | Liu et al. | Средняя юра | Формация Шаксимяо (Shaximiao Formation) |
(Сычуань) |
Цинодонт из семейства тритилодонтид. | ||
| Cifellilestes[171] | Gen. et sp. nov | Davis et al. | Верхняя юра (титон) | Формация Моррисон (Morrison Formation) |
( |
Морганукодон. Включает единственный вид — C. ciscoensis. | 
|
|
| Eoscansor[172] | Gen. et sp. nov | Lucas et al. | Карбон (миссисипий) | Формация Эль-Кобре-Каньон (El Cobre Canyon Formation) |
( |
Представитель семейства Varanopidae. Включает единственный вид — E. cobrensis. | 
|
|
| Euchambersia liuyudongi[173] | Sp. nov | Liu & Abdala | Верхняя пермь (вучапин) | Формация Наобаогоу (Naobaogou Formation) |
(Внутренняя Монголия) |
Тероцефал. |  |
|
| Guttigomphus[174] | Gen. et sp. nov | Rayner et al. | Скорее всего, средний триас (анизий) | Формация Бюргерсдорп (Burgersdorp Formation) |
(Восточно-Капская провинция) |
Трираходонтид. Включает единственный вид — G. avilionis. | 
|
|
| Kembawacela yajuwayeyi[175] | Sp. nov | Araújo et al. | Верхняя пермь (вучапин) | Чивета-Бедс (Chiweta Beds) | (Северная провинция) |
Дицинодонт из семейства Cistecephalidae. | 
|
|
| Koksharovia[176] | Gen. et sp. nov | Suchkova, Golubev & Shumov | Средняя пермь | Котельнич | ( |
Тероцефал из семейства Scylacosauridae. Включает единственный вид — K. grechovi. | 
|
|
| Lalieudorhynchus[177] | Gen. et sp. nov | Werneburg et al. | Средняя пермь (роуд/ворд — нижний кепт) | Формация Ла-Льед (La Lieude Formation) | Казеид. Включает единственный вид — L. gandi. | 
|
||
| Notictoides[178] | Gen. et sp. nov | Sidor, Kulik & Huttenlocker | Нижний триас | Формация Фремоу (Fremouw Formation) |
Тероцефал. Включает единственный вид — N. absens. |
|
[18] | |
| Phorcys[179] | Gen. et sp. nov | Kammerer & Rubidge | Средняя пермь (ворд — кепт) | Формация Абрахамскраль (Abrahamskraal Formation) |
Ранний горгонопс. Включает единственный вид — P. dubei. | 
|
||
| Tessellatia[180] | Gen. et sp. nov | Gaetano et al. | Верхний триас (норий) | Формация Лос-Колорадос (Los Colorados Formation) |
( |
Цинодонт клады Probainognathia. Включает единственный вид — T. bonapartei. |  |
{kind=link}
- Benoit et al. опубликовали исследование эволюции мозга, органов чувств и поведения немаммальных синапсид[181].
- Reisz, Scott & Modesto опубликовали исследование анатомии черепа Cotylorhynchus romeri[182].
- Bhat, Shelton & Chinsamy изучили костную гистологию различных таксонов дейноцефалов из средней перми ЮАР (Tapinocephalus Assemblage Zone, бассейн Кару), сделав выводы об их палеобиологии и палеоэкологии[183].
- Macungo et al. опубликовали исследование анатомии базикраниальной оси дицинодонтов надсемейства Emydopoidea. Исследователи обнаружили доказательства роющих адаптаций основания черепа (базикрания) у изученных таксонов, которые были интерпретированы как поддерживающие поведение рытья с поднятой головой, по крайней мере, для некоторых Cistecephalidae[184].
- Kammerer et al. описали новый материал Dicynodon angielczyki из Мозамбика (Metangula Graben) и Замбии (Luangwa Basin). Это первые экземпляры данного вида, обнаруженные за пределами Танзании (Ruhuhu Basin)[185].
- Liu сообщил о новом материале дицинодонтов Shaanbeikanemeyeria и Parakannemeyeria из среднетриасовой формации Эрмайинг (Ermaying Formation; Ordos Basin, Шэньси, Китай), который позволил переописать морфологию и таксономию этих таксонов[186].
- Sidor описал элементы частично сочленённого скелета небольшого горгонопса: правую подвздошную кость и кости задних конечностей, обнаруженные в отложениях верхнепермской формации Мадумабиса-Мадстоун (Madumabisa Mudstone Formation) в Замбии[187].
- Liu & Yang описали образец горгонопса из формации Утунгоу (Wutonggou Formation; Turpan Basin, Синьцзян, Китай). Открытие свидетельствует о том, что горгонопсы существовали в северной тёплой умеренной зоне около 253,3 миллионов лет назад, то есть примерно в то же время, что и другие последние известные представители этой группы, жившие на территориях нынешних России и ЮАР[188].
- Norton et al. провели исследование закономерностей смены зубов у позднепермского цинодонта Cynosaurus suppostus, изучив морфологию пяти образцов, которые, как предполагается, соответствуют разным онтогенетическим стадиям[189].
- Filippini, Abdala & Cassini оценили массы пяти видов циногнатий из триаса Аргентины (Andescynodon mendozensis, Pascualgnathus polanskii, Massetognathus pascuali, Cynognathus crateronotus и Exaeretodon), используя линейные измерения и длины окружностей их плечевых и бедренных костей[190].
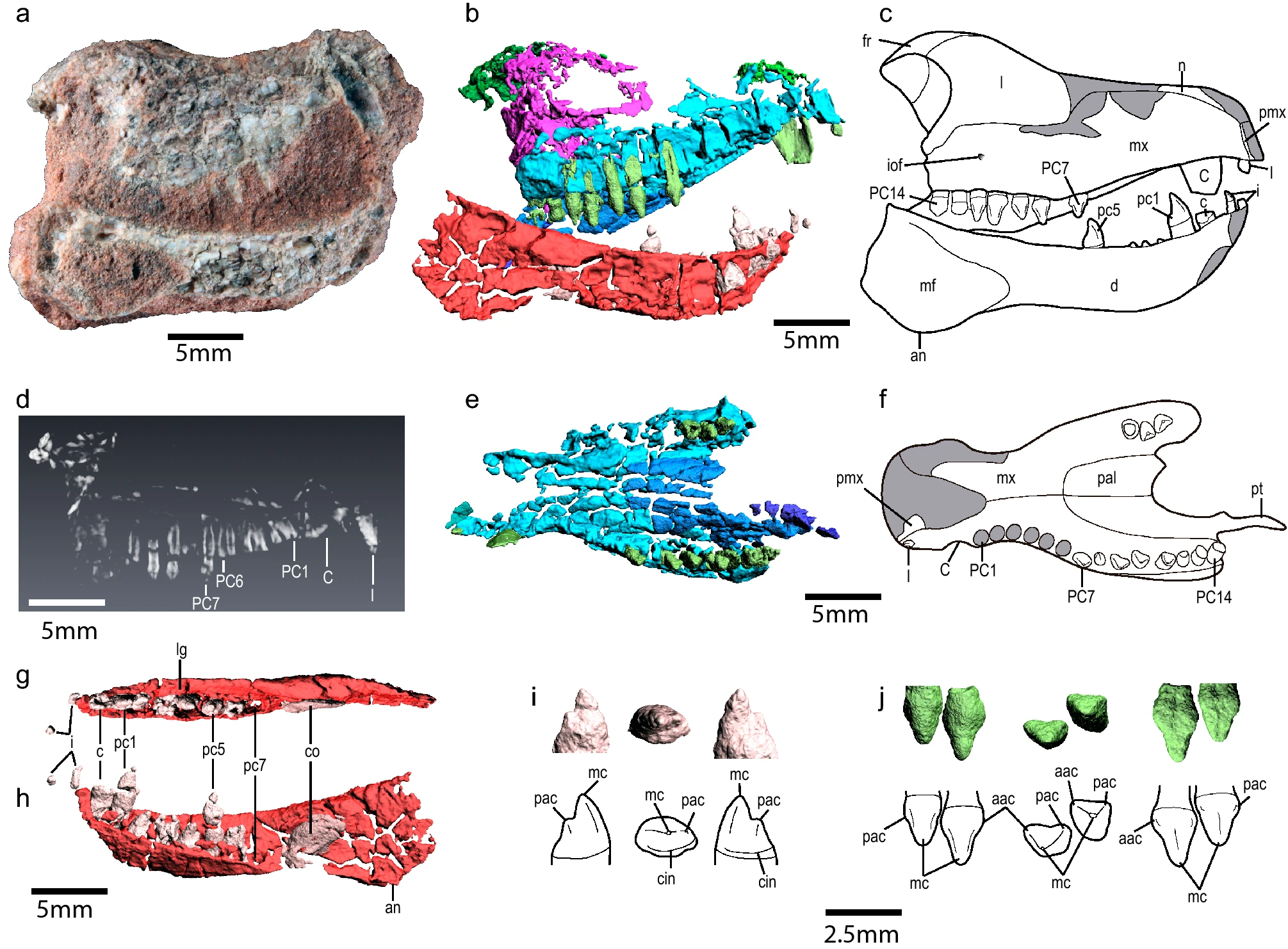
- Melo, Martinelli & Soares описали новый материал цинодонтов из верхнего триаса Бразилии (Santacruzodon Assemblage Zone, Santa Cruz Sequence, Santa Maria Supersequence): многочисленные остатки Santacruzodon hopsoni, позволившие уточнить анатомию этого вида, а также кости Chiniquodon sp.[191]
- Liu et al. описали новый материал тритилодонтида Lufengia из нижнеюрской формации Луфэн (Lufeng Formation; Юньнань, Китай), в котором интерпретировали остатки, ранее отнесённые к Dianzhongia, как принадлежащие более старой особи Lufengia[192].
- Luo et al. пересмотрели морфологию нижней челюсти и зубов маммалиаформа Hadrocodium wui[193].
- Исходя из того, что переход к эндотермии приводит к повышению температуры тела и росту подвижности, Araújo et al. разработали «индекс температуры и подвижности» (thermo-motility index, TMI), который вычисляется на основе морфометрических признаков костного лабиринта внутреннего уха. Используя расчёты TMI, исследователи пришли к выводу, что эндотермия резко развилась в течение позднего триаса у первых представителей клады Mammaliamorpha, тогда как все более базальные синапсиды были эктотермами[194][195].
Ссылки
Обзор палеонтологических открытий 2022 года; рассказывает доктор биологических наук Павел Петрович Скучас: